
UVC
Product Resources
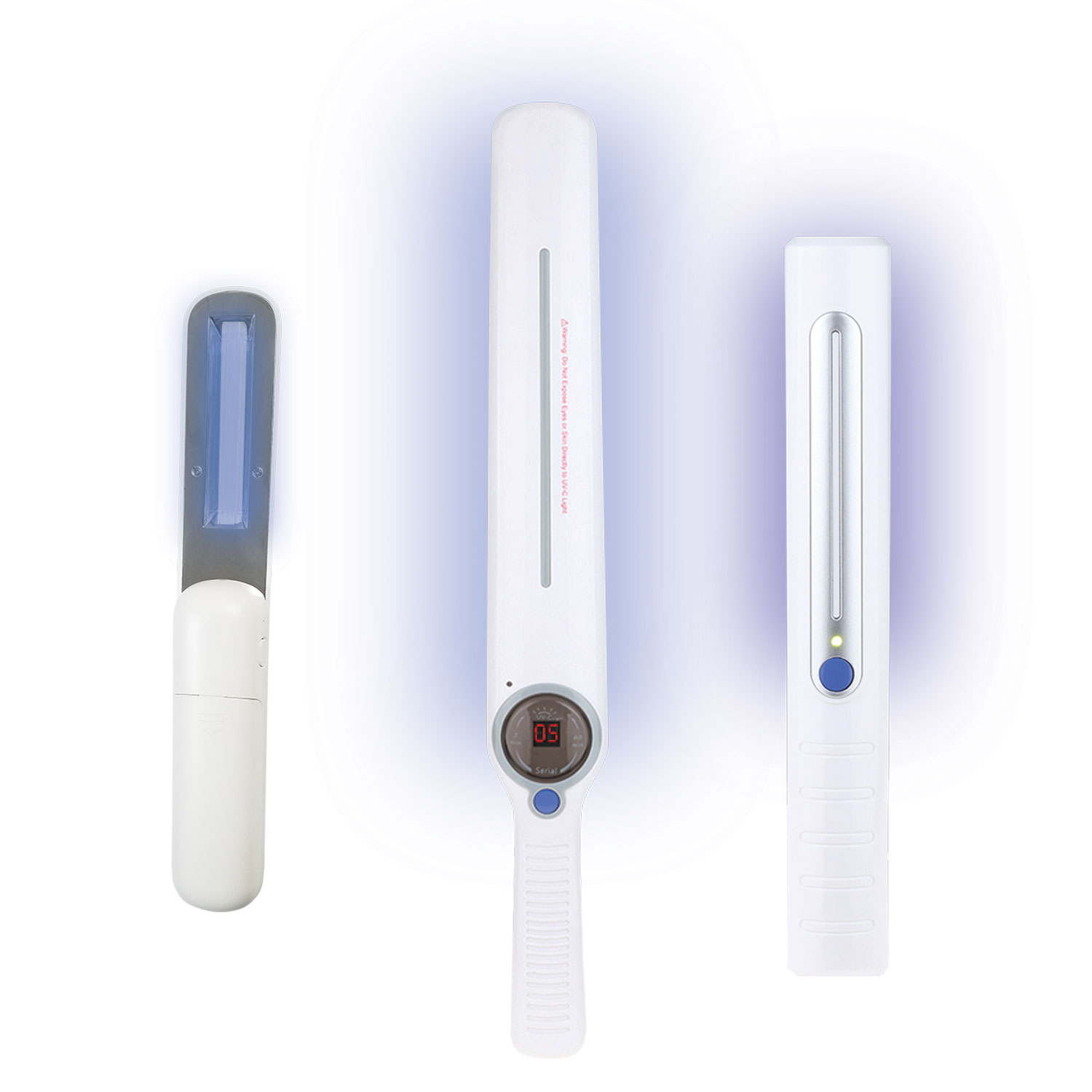
Rapid Handheld UVC Sanitizer Wands
Our powerful set of cordless wands will sterilize a surface within 25 seconds. Hover 1/2-2 inches above your desired area and let it work its magic. Both larger models have a safety feature to shut off when facing upward. The pocket-sized models fold.
- • Personal Use • Travel • Home • Dining Out • Small Office • Register

Commercial UVC Sterilizer
This effective and efficient sterilizer can disinfect a ~225 ft2 room in 15 minutes. With a range of up to ~650 ft2, this light has intelligent motion sensor safety features that deactivate the light if someone enters the area.
- • Hotel • School • Clinic • Convenience Store • Veterinary • Bank
UVC Sterilization Troffer
A permanent fixture to regularly sterilize a room, this troffer covers a 350-400 ft2 area at a height of 10-20 ft. It comes standard with remote control and hard-wired operation, with the option to add mobile app capability to manage lighting groups.
- • School • Office • Airport • Retail • Nursing Home • Drop Ceiling


UVC High-Powered Induction Lamp
Primarily used for air purification within HVAC systems, this intense ultraviolet lamp can maintain sterilization for an air volume of 70,500 ft3. Units like these can be controlled with a switch or wired to turn on automatically with other systems.
- • HVAC Air Purification • Water Treatment • Surface Decontamination
UVC Fully Automatic Sterilizer
Create a clean and safe experience for your clients with a fully automated UVC system. Intelligent programming allows this device to clean touch points on self-service machines after each and every visitor. No human intervention is required.
- • Gas Station Pump • Kiosk • Game Machine • Self Service Machine


UVC Intelligent ATM Sterilizer
This smart UVC system automatically sterilizes self-service machines in between visitors, and angles inwards to prevent light exposure in crowded areas. Sensors and programming allow it to run without human intervention.
- • ATM • Cabinet • Self Service Machine
Mobile UVC Sterilizer Units
Unfold the sterilizer arms to allow maximum exposure and clear a room of up to ~1,500 ft2. This light is perfect for facilities that require thorough residue-free cleaning. This product comes in two variations based on your requirements.
- • Hospital • Gymnasium • Warehouse • Retail • Mall • Church


Suspended UVC Sterilizer
Perfect for restaurants and industrial environments, this remote controlled UVC Sterilizer affects a ~400 ft2 area with 99.9% effectiveness against pathogens. That way you can keep a work area consistently safe and sanitized.
- • Restaurant • Casino • Community Center • Workshop • Nursing Home
UVC Sterilization Bag
The LED chips inside this bag emit both UVC and UVA light to sterilize with power greater than 20X the intensity of the sun. Plus, it only takes 5 minutes to disinfect. A gravity sensor in the lid prevents this from activating while it is open.
- • Salon • Barber Shop • Massage Parlor • Daycare • Home • Beauty


UVC Sterilizing Wireless Charger
This smart phone fast charging box utilizes UVC and Ozone to sterilize every crevice of your device. The aromatherapy oil diffuser inside adds a pleasing fragrance during disinfection. This is large enough to fit most mobile phones.
- • Home • Vehicle • Office • Lobby • Phone Store • Wholesale
UVC Educational Resources
Please Note:
UVC light can cause loss of vision and temporary acute redness or ulceration (sunburn) to exposed skin. To prevent exposure, do not operate the device in any way that allows UVC light to be visible during operation. Do not stare directly at the activated bulb or disinfecting target zone. Avoid touching a UVC bulb with bare hands. Fingerprints may result in reduced performance and can significantly shorten the lifespan of the lamp unless they are removed with alcohol. Please read all instruction manuals for UV product safety requirements before the operation.